--:-----
Триасовый период
09 июня’ 2015 | 07:57
INNA
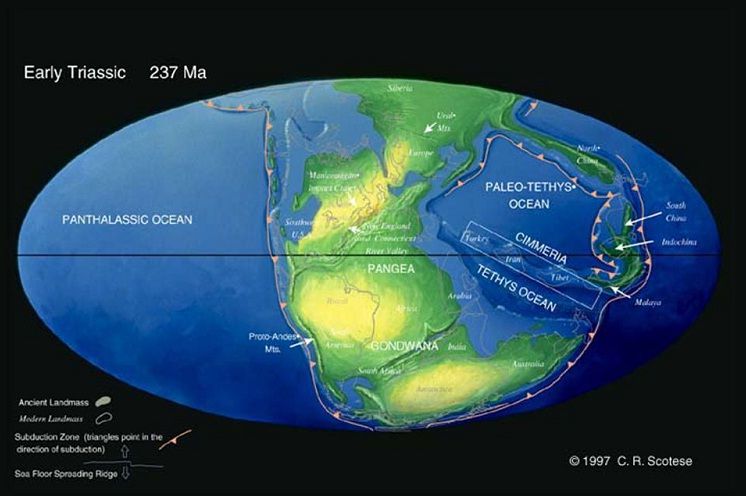
Триасовый геологический период, триас (250 - 199 млн. лет назад по другим данным 248 - 213 млн. лет) - длившийся приблизительно 50 млн. лет и являющийся первым периодом Мезозоя. Триасовый период подразделяют на три отдела: нижний триас (Скифский), средний триас (Ракушечный) и верхний триас (Кейперский).

Название «триасовый период» происходит от греч. trias (троица) в связи с четким трехчленным строением толщи отложений этого периода в северной Германии. В основании толщи залегают красноцветные песчаники, в середине – известняки, а вверху – красноцветные песчаники и глинистые сланцы. На протяжении триаса значительные территории Европы и Азии были заняты озерами и мелководными морями. Эпиконтинентальное море покрывало Западную Европу, причем его береговая линия прослеживается на территории Англии. В этом морском бассейне и накапливались вышеупомянутые стратотипические осадки. Песчаники, залегающие в нижней и верхней частях толщи, отчасти имеют континентальное происхождение. Другой триасовый морской бассейн проникал на территорию северной России и распространялся к югу по Уральскому прогибу.

Огромное море Тетис тогда покрывало примерно такую же территорию, как и в позднекаменноугольное и пермское время. В этом море накопилась мощная толща доломитовых известняков, которыми сложены Доломитовые Альпы северной Италии. На юге центральной Африки триасовый возраст имеет бoльшая часть верхней толщи континентальной серии кару. Эти горизонты известны обилием ископаемых остатков пресмыкающихся. В конце триаса на территории Колумбии, Венесуэлы и Аргентины образовались покровы алевритов и песков континентального генезиса.

До наступления триаса все материки существовали в виде единого гигантского суперматерика — Пангеи. С наступлением триаса Пангея начала постепенно раскалываться на Гондвану и Лавразию, начал образовываться Атлантический океан. В триасе сильно сокращаются площади внутриконтинентальных водоемов, развиваются пустынные ландшафты. К этому периоду относится Таврическая формация Крыма (верхний триас). Потепление климата вызывает высыхание многих внутренних морей. В оставшихся морях растёт уровень солёности.

Наряду с изменениями в распределении моря и суши, формированием новых горных массивов и вулканических областей, интенсивно проходила смена одних животных и растительных форм другими. Лишь немногие семейства перешли из палеозойской эры в мезозойскую. Первые 10 миллионов лет биосфера триасового периода мучительно оправлялась от катастрофы, вызвавшей самое большое вымирание в истории Земли, затем жизнь восстановилась в прежнем объеме и разнообразии.

Сезонное изменение температур триасового периода начало оказывать заметное влияние на растения и животных. Отдельные группы пресмыкающихся приспособились к холодным сезонам. Именно от этих групп, в триасе произошли млекопитающие, а несколько позднее — и птицы.

Похолодание в триасовом периоде было незначительным. Сильнее всего оно проявилось в северных широтах. На остальной территории было тепло. Поэтому пресмыкающиеся чувствовали себя в триасовом периоде достаточно хорошо. Разнообразнейшие их формы, с которыми мелкие млекопитающие еще не способны были конкурировать, расселились по всей поверхности Земли. Необычайному расцвету пресмыкающихся способствовала также богатая растительность триасового периода.

Растительный мир суши 1-й половины триасового периода во многом близок к верхнепермскому; в тропиках преобладают, ныне вымершие, птеридоспермы и древние хвойные, в умеренной над ними господствуют разнообразные палеофитные папоротники. На всех континентах широко распространены своеобразные плауновидные, специфические для триасового периода.

Основные мезофитные группы растений (диптериевые папоротники, саговники, беннеттиты, гинкговые, мезофитные хвойные) в значительном количестве появляются во 2-й половине триасового периода, но вплоть до конца триасового периода ещё велико значение древних групп. В морях триасового периода большую роль играли рифообразующие водоросли (Альпы).

В разное время триаса в разных группах насекомых происходили основные изменения. Например, сетчатокрылые виды в течение всего периода становились более разнообразными. В раннем триасе не было наземных жуков, хотя в перми они были широко распространены.

В среднем триасе стали расцветать полужесткокрылые, жесткокрылые (предположительно большинство - водные формы), стрекозы и подёнки. В позднем триасе появляется один из последних больших отрядов насекомых - двукрылые, а также перепончатокрылые.

Достоверно известно и доказано, что в конце триаса существовали прямокрылые; у самцов некоторых видов имелся звуковой аппарат на передних крыльях, с помощью которого они привлекали самок. В конце триаса вымерло одно из восьми семейств стрекоз.
На границе триаса и юры, синхронно с великим морским вымиранием, происходило и падение разнообразия насекомых, хотя основные изменения их состава произошли ранее, еще в позднем триасе.

Одна из групп триасовых рептилий - цинодонты. Их название переводится как "собачьи зубы". Они появились около 220 млн.л.н. Тело цинодонтов было покрыто шерстью, а крепкие челюсти и зубы идеально подходили для пережевывания мясной пищи. Ценодонты были революционной ветвью рептилий. Не смотря на то, что они откладывали яйца, малыши питались молоком матери. Впоследствии из цинодонтов разовьется новая ветвь животных, которых назовут млекопитающими. На протяжении последующих 160 миллионов лет, млекопитающим суждено находиться в тени динозавров. Только после вымирания динозавров, потомки цинодонтов смогут развиться и править землей.

В начале триасового периода появилась новая группа рептилий — архозавры, то есть "правящие рептилии". Архозавры были предками всех основных групп известных в мезозойскую эру. Потомками архозавров были все виды динозавров, крокодиломорфы, нотозавры, плиозавры и плезиозавры, ихтиозавры, плакодонты и птерозавры.

В последней трети триаса выделились две группы рептилий. Одни стали предками современных крокодилов. Другая группа развилась в динозавров.

Первые динозавры появились в последней трети триасового периода они принципиально отличались от прочих других видов рептилий строением костей таза. В отличии от лап рептилий, лапы которых были расставлены в стороны, лапы динозавров находились под туловищем. Благодаря этому отличию динозаврам было легче передвигаться. Динозавры ходили либо на двух, либо на четырех лапах. Некоторые динозавры были массивными и медленными, а другие передвигались быстро благодаря чему динозаврам было легче находить пищу и скрываться от врагов. Одними из первых динозавров были проворные целофизисы, крепкие герреразавры и гигантские по тем временам платеозавры.
Более совершенное строение скелета это всего лишь одна из причин такого успеха динозавров. Кроме этого кожа динозавров чешуйчатая и водоотталкивающая. Чешуя хорошо защищала динозавров от влаги и от хищников.

Яйца динозавров имели прочную скорлупу и поэтому процент выживания детенышей был очень высок.
Кроме динозавров, из архозавров, развилась еще одна группа рептилий принципиально отличающиеся от других. Эти рептилии приспособились к полету и назывались птерозавры. Первые птерозавры жили на берегах рек и питались насекомыми. В течение всей мезозойской эры птерозавры правили в воздухе.

Нотозавры были морскими хищниками, которые развились в триасовом периоде из сухопутных рептилий. Они имели длинное обтекаемое тело, тонкую шею и хвост и охотились исключительно на рыбу. Этот ископаемый скелет был найден в Швейцарии.

Аммониты, впервые появившиеся на Земле в девонский период, наиболее изученные и часто встречающиеся ископаемые морские животные. Они принадлежат к группе головоногих моллюсков и, стало быть, являются предшественниками современных осьминогов и кальмаров. Своего наивысшего расцвета, если иметь в виду их численность и разнообразие, аммониты достигли в пермский период. Затем 245 млн лет назад, в конце этого периода, они почти полностью исчезли при массовом вымирании. Но разделаться с аммонитами оказалось не так-то просто. Некоторые из них сумели дожить до триасового периода и благодаря своим склонностям к дальним странствиям по океану, вскоре вновь распространились по всему миру.

К середине мезозоя они достигли нового пика эволюционного процветания. Аммониты столь часто встречались в мезозойских морях, а их окаменелости в таком изобилии попадаются в горных породах той эпохи, что они сыграли весьма важную роль в разработке системы идентификации (отождествления) всех морских отложений мезозойской эры. И все же подобное процветание не могло быть вечным, и в конце мелового периода все аммониты внезапно исчезли с лица Земли — вместе со множеством других морских животных, включая белемнитов, плиозавров, ихтиозавров и плезиозавров.

У первых головоногих моллюсков, так называемых наутилоидей, были длинные раковины конической формы. Внутри раковин находились газовые камеры, разделенные перегородками. Наутилоидеи выработали весьма примитивный способ втягивания и выталкивания воды, причем выбрасываемую струю они использовали для создания своего рода реактивной тяги. За миллионы лет, прошедшие с тех пор, головоногие, включая аммонитов, усовершенствовали этот метод и сделали его своим главным способом передвижения. На протяжении всей палеозойской эры (570—225 млн лет назад) наутилоидеи были наиболее распространенными морскими хищниками. Потом появились новые, более высокоорганизованные головоногие, в том числе аммониты, у которых развилась раковина совсем иной формы — завитая, плоская и зачастую с довольно сложным рельефным узором.

Как и у современных наутилусов, раковина аммонита была разбита на ряд внутренних камер. Каждая из них отделялась от соседней перегородкой. Само животное помещалось в самой последней из новообразованных камер. Места, в которых перегородки присоединялись к раковине, часто хорошо видны на ископаемых останках аммонитов. Их называют шовными линиями. Такие линии образуют на раковинах аммонитов, живших в юрский и меловой периоды, довольно сложные узоры. Ученые используют эти узоры для классификации огромного количества ископаемых аммонитов, обнаруженных за годы исследований.

Раковина аммонита служила как бы балластным (стабилизирующим) механизмом, подобно тому как нынешние наутилусы используют собственные многокамерные раковины. Животное заполняло свободные камеры водой и вновь их опорожняло при помощи особого органа — его называют сифоном. Закачивая и выкачивая воду, аммонит регулировал плавучесть собственного тела, как это делают подводные лодки. Когда животное хотело поглубже погрузиться, оно наполняло свои "балластные отсеки" водой. Когда же оно желало всплыть на поверхность или просто подняться выше, — опорожняло их.
Невероятное изобилие ископаемых останков аммонитов оказалось несколько обманчивым, заставив ученых в свое время сделать не совсем верные выводы об их распространении в прошлые эпохи. До самого последнего времени специалисты полагали, что аммониты населяли все первобытные моря без исключения. Однако затем было установлено, что отнюдь не все их разновидности обитали по всему земному шару. У разных видов были различные ареалы, что, по всей вероятности, было связано с рядом физических факторов, например с температурой воды или с процентным содержанием в пей соли (соленостью). Смещение материков в конце мелового периода привело к грандиозным климатическим изменениям и общей трансформации (преобразованию) природной среды, затронувшей и Мировой океан. Вероятно, аммониты на сей раз не сумели приноровиться к новым условиям жизни и полностью вымерли.
Комментарии:
Нет комментариев